Des cellules qui migrent collectivement en distinguant leur gauche de leur droite
Des physiciens ont étudié comment la présence d’une paroi induit un mouvement collectif dans un tissu cellulaire. La forte activité de ces cellules, leur forme allongée et une asymétrie microscopique concourent à engendrer ces déplacements près d’une paroi, favorisant ainsi leur migration collective via un mécanisme purement physique.
En biologie, la question du déplacement collectif des structures pluricellulaires est très importante, car la mobilité de ces dernières conditionne leur possibilité de développement voire leur survie. Alors que ces déplacements cellulaires coordonnés ont longtemps été imputés à des mécanismes de coordination d’origine biologique ou chimique, les physiciens du vivant ont montré récemment que leur émergence est aussi un phénomène physique opérant à des échelles de longueur sensiblement plus grandes que la taille d’une cellule unique.
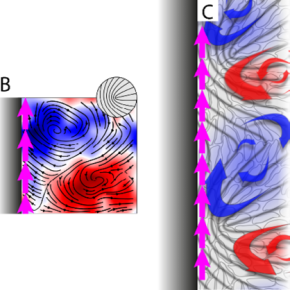
Une collaboration internationale réunissant des scientifiques du Laboratoire physico-chimie Curie (PCC, CNRS / Institut Curie / Sorbonne Université), de l’Université de Leiden aux Pays Bas et de l’Université Ben Gourion en Israël s’est ainsi récemment intéressés à la migration collective de cellules de fibrosarcome, un cancer particulièrement agressif. Ces cellules, en forme de fuseau, ont été mises en culture sur des bandes dont la largeur est grande devant une taille cellulaire et délimitées latéralement par deux surfaces antiadhésives. Les cellules prolifèrent et colonisent rapidement la bande (aux échelles biologiques, soit environ 24 heures) jusqu’à former une monocouche, parcourue de mouvements internes de réorganisation, désordonnés et chaotiques. À distance des bords, ce désordre n’induit pas de déplacement net. En revanche les chercheurs ont remarqué que lorsque les cellules sont très proches des bords de la bande, elles migrent systématiquement le long de ceux-ci (Figure C, le mouvement y est dirigé suivant les flèches magenta). Ces courants de bords sont toujours dirigés dans le même sens (le mouvement global est systématiquement vers la droite lorsqu’on regarde la frontière depuis la monocouche). Cette migration est induite par la présence d’une frontière et non par le confinement, la zone de tissu concernée par cette migration -quelques tailles cellulaires tout au plus- restant beaucoup plus petite que la largeur de la bande. Ainsi, la simple présence d’une paroi est susceptible d’organiser l’activité des cellules et de faire émerger ce déplacement d’ensemble. Ce phénomène est à rapprocher d’observations in vivo chez la souris montrant des cellules cancéreuses migrant collectivement le long des interfaces avec d’autres tissus.
Pour comprendre ce courant cellulaire émergent qui semble briser les symétries du système, les physiciens ont modélisé la monocouche de cellules comme un fluide continu nématique (ce qui veut dire que la forme des cellules en bâtonnets lui confère une orientation locale), et actif (les cellules transformant les nutriments de leur environnement en forces mécaniques donc en écoulements). Le caractère nématique du fluide produit des singularités dans le champ d’orientation (des défauts topologiques) que l’activité cellulaire met spontanément en mouvement via des paires de tourbillons. Quand ils rencontrent une paroi (une des bandes antiadhésives), ces défauts ne peuvent plus avancer et on s’attendrait à ce qu’ils s’auto-organisent de manière stable perpendiculairement à la paroi. Mais c’est sans compter sur la chiralité cellulaire, une asymétrie dynamique qui tire son origine de l’asymétrie des filaments constituant le squelette de la cellule. Dans la monocouche, loin des bords, cette chiralité conduit à une légère inclinaison de la vitesse des défauts par rapport à leur axe (Figure A). Près d’une frontière physique, cet effet se traduit en une inclinaison de l’axe des paires de tourbillons par rapport à la direction de la bande (Figure B), créant ainsi un écoulement net dont la direction est fixée par le signe de la chiralité. Ce courant de bord a de plus pour conséquence de faire pivoter l’axe des défauts, ce qui renforce encore l’écoulement (Figure C).
Il s’agit d’une situation dans laquelle le biais dû à la chiralité va plus loin que de fixer la direction du mouvement puisqu’il est ici directement responsable de l’existence même de ces courants de bord. Quantitativement, la contrainte chirale n’a pas besoin d’être importante (dans ce travail, elle ne s’élève qu’à environ 3% de la contrainte totale) mais le déséquilibre qu’elle engendre est suffisant pour produire ces courants. Ce dernier résultat laisse penser que ce type de mécanisme pourrait être à l’origine de déplacements cellulaires collectifs dans d’autres situations biologiques.
Ces résultats sont publiés dans la revue Physical Review X.
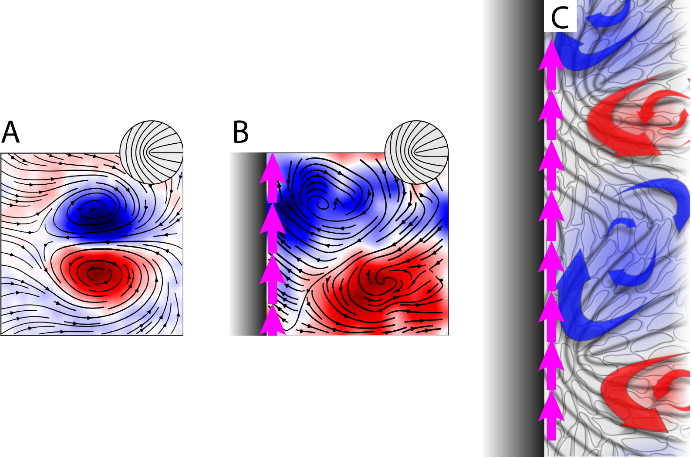
A- Loin des bords de la bande adhésive, les défauts topologiques s’accompagnent de paires de tourbillons. La couleur code pour la direction de ces tourbillons (bleu dans le sens horaire, rouge dans le sens anti-horaire). Les lignes sont les lignes du champ de vitesse. La vitesse du défaut fait alors un angle faible avec sa direction (encart).
B- Près du bord (en noir) cette paire de tourbillons pivote par rapport à la direction de la bande et engendre ainsi un écoulement net (flèches magenta). Ce courant de bord provoque l’inclinaison des défauts eux-mêmes (encart), ce qui tend à le renforcer.
C- Vue d’artiste illustrant le mécanisme donnant lieu à un courant de bord (flèches magenta) dans un tel système chiral actif. Les lignes grises représentent ici le champ d’orientation des cellules. Les flèches bleues et rouges correspondent aux écoulements dans les tourbillons. Les dimensions des figures A et B sont de 250 x 250 µm².
Références
Chiral Edge Current in Nematic Cell Monolayers, V. Yashunsky, D. J. G. Pearce, C. Blanch-Mercader, F. Ascione, P. Silberzan, and L. Giomi, Phys. Rev. X, publié le 14/11/2022.
DOI : 10.1103/PhysRevX.12.041017
Archives ouverte HAL